Skip to main content
Primary Research Projects
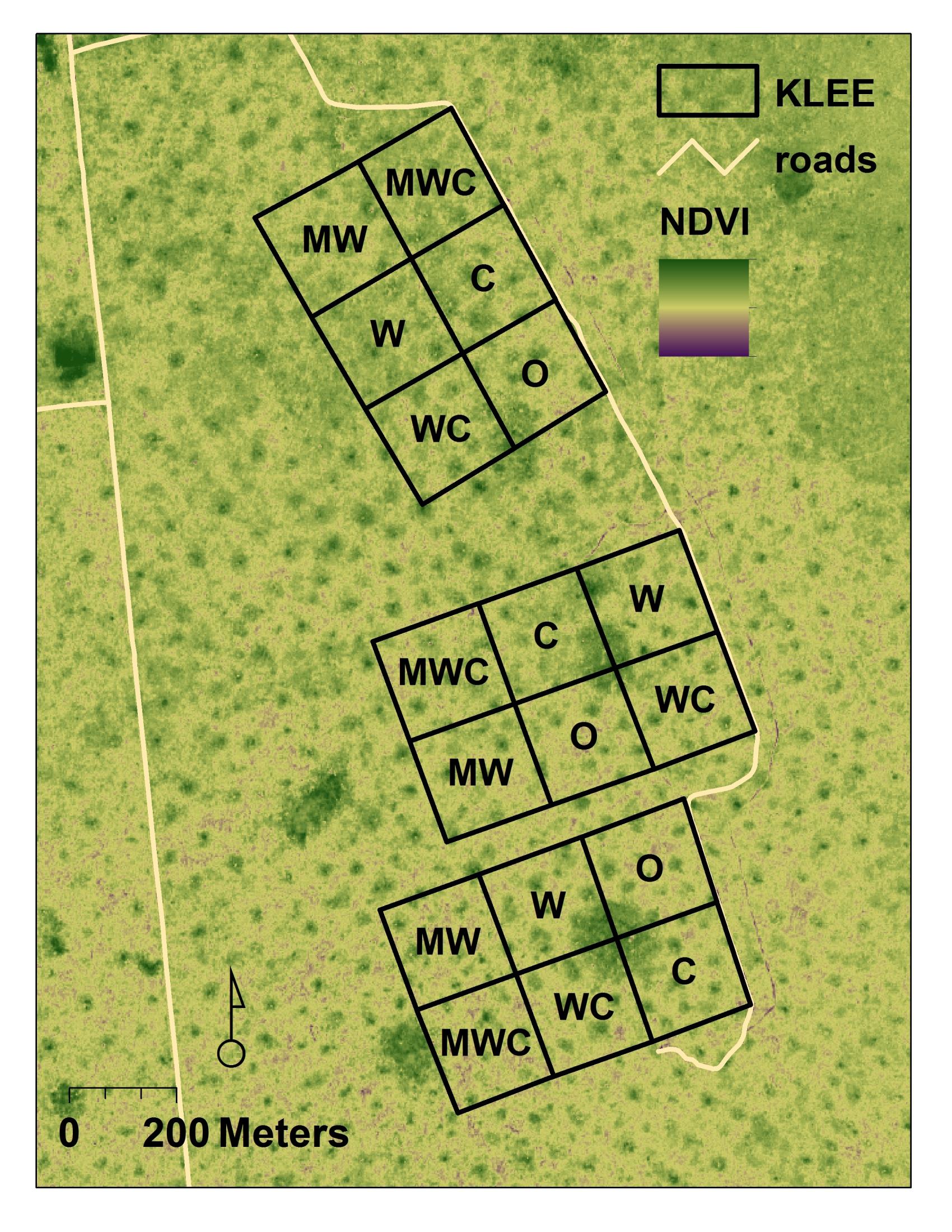
KLEE is a long-term, large-scale herbivore exclusion experiment in Laikipia, Kenya funded by the National Science Foundation and the National Geographic Society. On arid and semi-arid range lands worldwide, large mammal diversity is in decline, and the centuries-old coexistence between livestock and native plant and animal biodiversity is threatened. Much of this land suffers from size-specific wildlife loss and degradation caused by changes in land management. Although there has been considerable research on the effects of individual wild and domestic herbivores on rangeland grassland ecosystems, there has been relatively little research on the separate and combined effects of different herbivore guilds on each other and on the vegetation they share. As the value of biodiversity climbs, and the profitability of livestock production declines, we are seeking more innovative ways to manage, conserve, and restore these landscapes.
In 1995, we established 18 four-hectare exclosures that allow herbivory by six different combinations of cattle, wildlife, and mega-herbivores (elephants and giraffes). KLEE is a controlled, replicated laboratory for the study of the effects of wildlife and livestock on each other and on the ecosystem they share. This experiment can be thought of as representing a) different land-use strategies, and b) different extinction scenarios. We have added additional experimental layers since, including two sets of controlled burns, tree thinning, "over-grazing" treatments, and the effects of mobile cattle bomas (corrals). Our collaborative team is monitoring soil, plant, invertebrate, and vertebrate responses to these experimental treatments (see KLEE Publications). These exclosures also allow us to address fundamental issues in ecology, including compensation, competition, and facilitation among herbivores, and ecosystem function, stability and resilience in the face of multiple stressors.
We are increasingly interested in the interactive effects on ecosystem resilience of multiple ecosystem drivers, including those associated with global change, which KLEE is uniquely able to address. KLEE has become the most productive field experiment ever carried out on the continent of Africa, with more than 128 peer-reviewed publications (174 from the overall KLEE project).
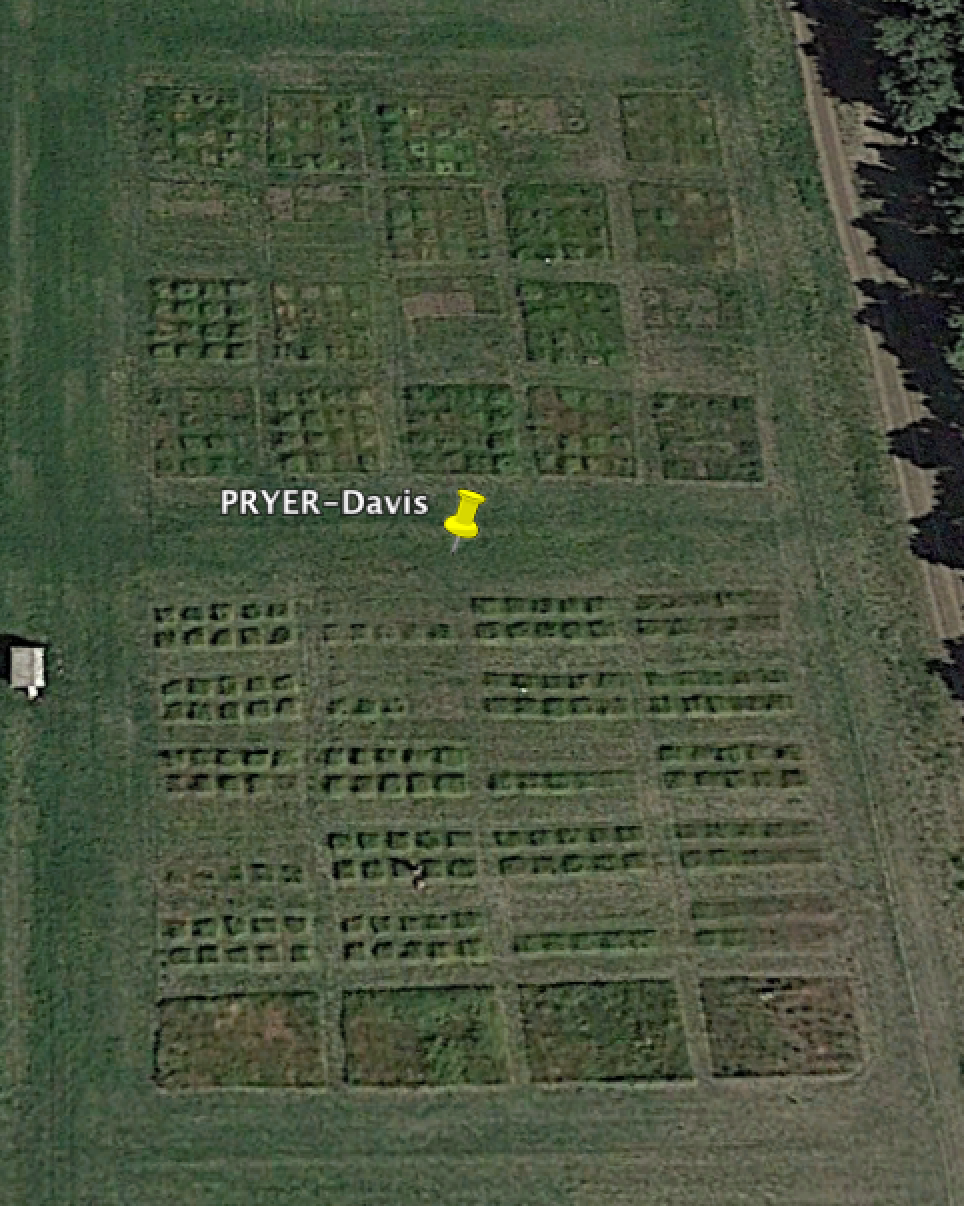
Contingency in community assembly: priority and year effects in restoration (PRYER)
Two alternative but potentially complementary paradigms shed light on the conceptual basis of community ecology for ecological restoration. Succession theory suggests that ecological restoration is the task of assisting, reestablishing, or accelerating the natural process of succession toward a desired (climax) state. Some versions of assembly theory suggest that there may be multiple alternative ecological states that can be derived from a given species pool, making restoration efforts ambiguous and difficult. At the heart of these models is the idea of priority effects, in which variation in early arrival by different species changes the course of community development through preemption of space or other limiting resources.
Supported the National Science Foundation, we examined the strength of such priority effects and possible mechanisms to encourage or suppress them in restoration settings, often in the context of California grassland communities. In particular, we tested both temporal priority effects (planting less dominant species a year earlier than more dominant species) and spatial priority effects (planting in small-sale monocultures versus seed mixes). But more than that, we examined a related form of contingency- year effects, in which year of establishment (or restoration) has profound effects on the trajectory of community development. We established these multiple priority experiments identically over five different years, and three California grassland restoration sites. In additional to documenting strong priority effects, we have shown that these experiments are profoundly affected by both year effects and site effects, as well as plant life history (annuals vs. perennials).
The maintenance of biodiversity in a model system: Ant Coexistence and Competition in Africa (ACACIA)
With Todd Palmer (University of Florida), Maureen Stanton (UC Davis) and Dan Doak (University of Wyoming), we began a research project in 1997 that gets at the conceptual heart of biodiversity conservation. One of the central questions in ecology is, “Why are there so many species?” The success of conservation and restoration projects is measured most fundamentally as species richness, and yet we still have only a rudimentary notion of what limits species diversity within and among communities. Our ability to restore and maintain biodiversity is limited by our understanding of how nature itself maintains species diversity. We studied the mechanisms of species co-existence in a system of acacias and their symbiotic ants uniquely suited to experimental, descriptive, and modeling approaches- the four ant species that inhabit Acacia drepanolobium of Laikipia, Kenya.
This research then expanded to descriptive and experimental examinations of the ecology and evolution of mutualisms in this system, including the benefits to trees of sequentially multiple ant partners and the examination of mutualisms as mutual parastisims. Most recently, this research group (including Corinna Riginos and Partick Milligan, and now independent of the Young Lab), has examined the devastating effects of the invasive big-headed ants (Pheidole megacephala) on populations of Acacia drepanolobium via the loss of their symbiotic ants.
Additional Research Projects
Critical issues in ecological restoration
Our lab has engaged in several projects designed to inform more effective ecological restoration, including:
Testing the cost-effectiveness of various planting techniques in restoration settings.
Exploring the role of fire in ecological restoration, conservation, and management, in African savanna rangelands and California prairies, oak woodlands, and coniferous forests.
Our California grassland restoration research has been supported by NSF and by CalFed through Audubon.
The evolution of semelparity (and plant population biology)
Some of my more theoretical, early research was on life history evolution and plant population biology. I particular interest has been the evolution of semelparity– the life history characterized by a single, massive, fatal reproductive episode. Organisms exhibiting semelparity range from the spectacular (certain salmon, bamboos, cicadas) to the mundane (many invertebrates), and include all of our (annual) grain crops. Biologists have long pondered how evolution could favor death after first reproduction. A key clue is in the observation that semeplarous organisms produce more offspring (or seeds) in their single reproductive event than do closely related iteroparous species in any one of theirs. My past research on Mount Kenya Lobelias and later work with Peter Lescia on Arabis fecunda suggest that a simple mathematical model explains the demographic conditions under which this increase in reproductive output more than compensates for the loss of potential future reproduction. I continue to dabble at the edges of these questions.
Induced defense
My interest in induced defense was first piqued over 40 years ago by looking at acacia spines near my house in Naro Moru, Kenya, and noticing that spines out of reach of local herbivores (goats) were significantly shorter than spines lower in the tree's canopy (Young 1987). Later I extended this work to acacia spines and mutualistic ants induced by giraffes (Milewski et al. 1992, Madden & Young 1992), and using experimental spine removal to demonstrate that these spines were in fact effective defenses again mammalian herbivory (Milewski et al. 1992). The KLEE experiment provided a controlled replicated test of the effects of different-sized herbivores on induced defense in Acacia drepanolobium. Spines produced during five years of protection from large mammalian herbivores were significantly shorter (Young & Okello 1998). In plots excluding all large herbivore (or all but cattle, who do not feed on A. drepanolobium), spines were far shorter at all branch heights. In plots excluding only mega-herbivores, spines were shorter only on branches higher than two meters (out of reach of other wildlife). Spines on branches more than 4m above the ground are always short, regardless exclusion treatment. Huntzinger et al. (2004) showed similar relaxation of rewards to mutualistic ants, in the form of fewer ants domatia (swollen thorns) and fewer extra-floral nectaries. This was associated with a shift in the acacia ant community from better protectors of acacias to indifferent ant species. This had profound implications for A. drepanolobium population dynamics (Palmer et al. 2008). Patterns of chemical defense (tannins) in response to KLEE herbivores treatments are complex (Ward & Young 2002). Simulated herbivory experiments showed that spine lengths can be rapidly re-induced (Gadd et al. 2001, Young et al. 2003). This pattern of slow relaxation and rapid induction may reflect differences in signal reliability: the absence of herbivory on a given branch in a given year (or even a few years) may not reliably predict the actual absence of herbivores, but the experience of herbivory is likely to be a reliable predictor of the presence of herbivores. Similarly, the relaxation of plant defenses may initially put plants at risk if herbivores are suddenly re-introduced (Young et al. 2021).

